Scientific background and current status of research
Sustainable intensification of agriculture and agroforestry
Agricultural production has increased remarkably over the past four decades, reaching historically high values, and is currently able to feed a global population of more than 7 billion people (Tilman et al. 2002). However, the agricultural practices that have been so successful in increasing global food supply have also resulted in unintended, negative impacts on the environment and on ecosystem services, emphasizing the need for more sustainable agricultural methods. Since a few decades people are aware of the need to increase the global agricultural production with better environmental protection (ŌĆśsustainable intensification of agricultureŌĆÖ) as shown by several review articles (Matson et al. 1997, Cassman 1999, Tilman et al. 2002). However, at present intensification of agriculture is still mainly attained through high fertilizer, water and pesticides use and new crop strains, and does not try to take advantage of ecological interactions within agricultural systems, which are crucial to secure high-productivity agriculture in the future (Robertson and Swinton 2005).
In natural ecosystems, the activity of a highly diverse community of soil microorganisms and invertebrates regulates key ecosystem properties such as decomposition, soil nutrient cycling and soil structure that directly affect productivity, without negative impacts on the environment. In agricultural systems, the composition and activity of the soil community differ markedly from natural ecosystems (Roesch et al. 2007) which may severely reduce the biological regulation of decomposition and nutrient availability in soils. In intensive agriculture this decoupling from biological activity is substituted by the use of mineral fertilizers and mechanized tillage. However, the addition of relatively mobile mineral nutrients which are not well synchronized with crop demand, and the inability of the soil biological community to retain excess nutrients, have led to leaching losses to surface or groundwater and trace gases emission to the atmosphere (Matson et al. 1997).
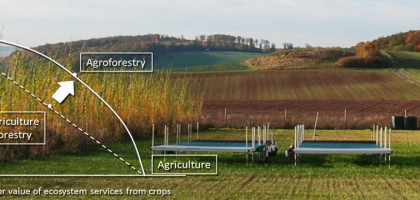
Sustainable intensification of agriculture can be attained if land use systems are designed that combine modern ecological knowledge of the functioning of ecosystems with traditional knowledge on farming. This awareness has stimulated renewed interest in agroforestry since there are several reasons why agroforestry systems are better able to mimic the functioning of natural ecosystems than monoculture crops. In general, growing trees with crops will be beneficial when the trees are able to acquire resources (water, light or nutrients) that the crop would otherwise not acquire, but will not be beneficial if the trees and crops are competing for the same resources (Cannell et al. 1996, Devkota et al. 2009). Agroforestry systems can have higher water use efficiency than annual crops since trees in agroforestry systems can utilize deep water outside the rooting zone of annual crops and outside the crop growing season. However, whether agroforestry systems can influence overall water consumption by reducing wind speed and affect infiltration rates at the field scale is still debated (Herbst et al. 2007). Furthermore, studies have shown that agroforestry systems can make more efficient use of available nutrients since deep tree roots can provide a ŌĆśsafety netŌĆÖ against nutrient leaching (Lehmann et al. 1998), supply nutrients from deep layers that are normally considered beyond the reach of crops (Dechert et al. 2005), and take up nutrients at times when crop demand is low. Other improved benefits that are often ascribed to the incorporation of trees in agroecosystems include stabilized or increased soil organic matter levels (Oelber-mann et al. 2004), improved aggregates stability, and stimulated extracellular enzyme activities (Udawatta et al. 2008).  Agroforestry systems in developed countries have only very rarely been analyzed in agricultural economics. Due to a growing interest in this land use system, some preliminary economic analyses have been conducted (Musshoff 2012). In Europe it has been shown that growing trees and crops in agroforestry systems generated a higher value of ecosystem services than growing them separately (Graves et al. 2007), see also Fig.1. Also in Germany, inclusion of tree components in agroecosystems is an approach that deserves thorough evaluation since there are no obvious reasons why agroforestry systems would not be equally successful, especially on ŌĆśmarginal sitesŌĆÖ and sites with high risk of nitrate leaching or erosion. Furthermore, agroforestry systems as an environmentally friendly land use have potential in marketing strategies which can enhance the image of an agricultural enterprise or a region (Reeg 2011).
Agroforestry systems in developed countries have only very rarely been analyzed in agricultural economics. Due to a growing interest in this land use system, some preliminary economic analyses have been conducted (Musshoff 2012). In Europe it has been shown that growing trees and crops in agroforestry systems generated a higher value of ecosystem services than growing them separately (Graves et al. 2007), see also Fig.1. Also in Germany, inclusion of tree components in agroecosystems is an approach that deserves thorough evaluation since there are no obvious reasons why agroforestry systems would not be equally successful, especially on ŌĆśmarginal sitesŌĆÖ and sites with high risk of nitrate leaching or erosion. Furthermore, agroforestry systems as an environmentally friendly land use have potential in marketing strategies which can enhance the image of an agricultural enterprise or a region (Reeg 2011).
Nutrient response efficiencies- four steps for higher productivity
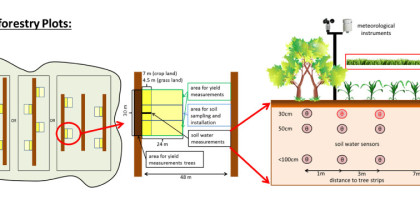
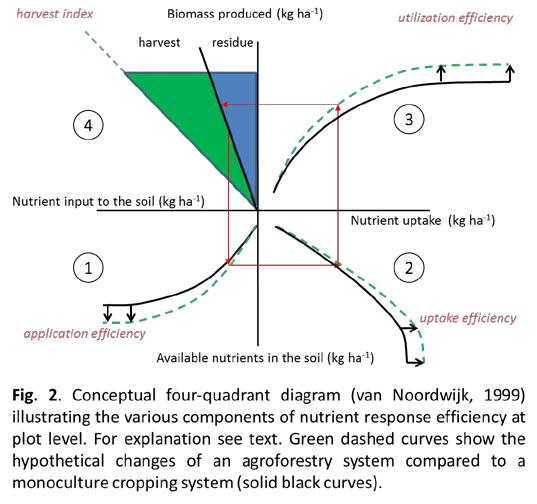 Traditional dose-response curves for fertilizer-yield relations (Cerrato and Blackmer 1990) often ignore that nutrient response efficiencies have various components as illustrated in Fig. 2 (Noordwijk 1999). Quadrant 1 at the lower left of Fig. 2 shows the relation between inputs of nutrients to the soil (e.g. in the form of crop residues, fertilizers) and the nutrients available in the soil during the growing season. This relation, the application efficiency, depends on inherent soil fertility, residue quality and quantity, timing, priming effects, placement, soil microbial processes (e.g. N-immobilization) and leaching. Quadrant 2 shows the relation between available nutrients and nutrient uptake by crops. The main drivers of this uptake efficiency are water uptake by roots, microbial biomass turn-over, and enzyme activities. Quadrant 3 shows the relation between nutrient uptake and biomass production. This utilization or use efficiency depends on factors such as weather conditions, cultivars, pests and competition. Finally, Quadrant 4 displays the ŌĆśharvest indexŌĆÖ, the ratio of harvest to biomass produced. Using these ideas, it was shown that in temperate grasslands nitrogen (N) response efficiencies depended on management and were mainly driven by N uptake efficiency rather than N use efficiencies (Keuter et al. 2013). Furthermore, it was shown at the same sites that microbial N immobilization rather than plant N uptake reduced N losses (Hoeft et al. 2014). It has frequently been observed that management practices, influencing the quantity and quality of plant residues, also influence the activity and diversity of soil organisms (Fliessbach et al. 2007). However, only very few studies exist for soils under agroforestry (Lagomarsino et al. 2011) and none was conducted in Germany. Labelling of plant residues in combination with analysis of microbial residues will give insights into the processes controlling nutrient application, uptake and utilization efficiencies and is thus essential to link soil biological functions to the efficient use of nutrients. Earthworms, a key component of soil fau-na that affect nutrient cycling through their feeding and burrowing activities, are also strongly influenced by management (Metzke et al. 2007). How their abundance and diversity links to nutrient dynamics in agroforestry systems is presently unknown. Whether agroforestry systems in Germany are indeed more efficient in nutrient cycling than conventional systems is also currently unknown. Since trees in agroforestry systems have the potential to acquire nutrients that crops would otherwise not acquire, this would potentially increase the ecosystemŌĆÖs application efficiency and uptake efficiency making the four-quadrant diagram a useful concept to evaluate agroforestry systems (Fig. 2).
Traditional dose-response curves for fertilizer-yield relations (Cerrato and Blackmer 1990) often ignore that nutrient response efficiencies have various components as illustrated in Fig. 2 (Noordwijk 1999). Quadrant 1 at the lower left of Fig. 2 shows the relation between inputs of nutrients to the soil (e.g. in the form of crop residues, fertilizers) and the nutrients available in the soil during the growing season. This relation, the application efficiency, depends on inherent soil fertility, residue quality and quantity, timing, priming effects, placement, soil microbial processes (e.g. N-immobilization) and leaching. Quadrant 2 shows the relation between available nutrients and nutrient uptake by crops. The main drivers of this uptake efficiency are water uptake by roots, microbial biomass turn-over, and enzyme activities. Quadrant 3 shows the relation between nutrient uptake and biomass production. This utilization or use efficiency depends on factors such as weather conditions, cultivars, pests and competition. Finally, Quadrant 4 displays the ŌĆśharvest indexŌĆÖ, the ratio of harvest to biomass produced. Using these ideas, it was shown that in temperate grasslands nitrogen (N) response efficiencies depended on management and were mainly driven by N uptake efficiency rather than N use efficiencies (Keuter et al. 2013). Furthermore, it was shown at the same sites that microbial N immobilization rather than plant N uptake reduced N losses (Hoeft et al. 2014). It has frequently been observed that management practices, influencing the quantity and quality of plant residues, also influence the activity and diversity of soil organisms (Fliessbach et al. 2007). However, only very few studies exist for soils under agroforestry (Lagomarsino et al. 2011) and none was conducted in Germany. Labelling of plant residues in combination with analysis of microbial residues will give insights into the processes controlling nutrient application, uptake and utilization efficiencies and is thus essential to link soil biological functions to the efficient use of nutrients. Earthworms, a key component of soil fau-na that affect nutrient cycling through their feeding and burrowing activities, are also strongly influenced by management (Metzke et al. 2007). How their abundance and diversity links to nutrient dynamics in agroforestry systems is presently unknown. Whether agroforestry systems in Germany are indeed more efficient in nutrient cycling than conventional systems is also currently unknown. Since trees in agroforestry systems have the potential to acquire nutrients that crops would otherwise not acquire, this would potentially increase the ecosystemŌĆÖs application efficiency and uptake efficiency making the four-quadrant diagram a useful concept to evaluate agroforestry systems (Fig. 2).
Water use efficiencies
Water use efficiency (WUE) expresses the biomass or grain yield produced per unit of water used by the land use system (Hatfield et al. 2001). Water use of crops and woodlands in the same region can vary significantly, particularly with respect to partitioning of evapotranspiration into transpiration, soil evaporation and interception evaporation (H├Črmann et al. 2008). Water uptake by roots is governed by water availability, root biomass and the spatial distribution of roots, but also by rhizosphere processes. In particular, mucilage exudation may play a key role in attenuating drought stress by keeping the rhizosphere wet and helping roots to take up water from drying soils (Carminati and Vetterlein 2013). For agroforestry systems it has been suggested that deep roots of trees can bring additional water to the top soil via hydraulic lift, providing a safety reservoir of water for shallow-rooted crops (Caldwell and Richards 1989). When evaluating the water balance of agricultural systems, several components (run-off, soil evaporation and drainage) do not contribute to crop productivity. Although there is evidence that agroforestry has the potential to increase WUE by reducing these unproductive components (Ong et al. 2002), it is presently unclear and scientifically very challenging to evaluate whether agroforestry systems in Germany have higher water use efficiencies than conventional agriculture.